����ʱ�䣺�������ࣺũҵ���������1��
ժ Ҫ�� ժ Ҫ ��Ŀǰ GenBank ���ݿ��¼ 167 ��ֱ��Ŀ����ȫ��������������У��漰����Ŀ 9 ���ܿ� 22 ���� 99 �����֣����Ŀ 7 ���ܿ� 12 ���� 68 �����֡��ڴ˻����ϣ����ķ�����ֱ��Ŀ���������������Ļ���������������������ȫ��������ֱ��Ŀ����ϵͳ�����о��ϵ�
����ժ��Ҫ ��Ŀǰ GenBank ���ݿ��¼ 167 ��ֱ��Ŀ����ȫ��������������У��漰����Ŀ 9 ���ܿ� 22 ���� 99 �����֣����Ŀ 7 ���ܿ� 12 ���� 68 �����֡��ڴ˻����ϣ����ķ�����ֱ��Ŀ���������������Ļ���������������������ȫ��������ֱ��Ŀ����ϵͳ�����о��ϵ�Ӧ�� ;ͬʱ����������ȫ�����������ؽ���ֱ��Ŀ�����ϵͳ������ϵ����Ҫ������� ��(1) ֱ��Ŀ������� 8 ��������������������ͣ����� trnK-trnD ��������������ڻ��ܿ��У�trnN-trnS-trnE ������������������ܿ��У�trnM-trnI-(-trnQ) �����������������Ҷ�x�ǿ��� ;(2) ֱ��Ŀ����ȫ�����������ļ����ɾ������Ե� AT ƫ���� ;(3) ��ͬ�ĵ����ʱ��������ֱ��Ŀ�����еĽ������ʲ�ͬ ;(4) ֧��ֱ��Ŀ�Լ����Ŀ�ͻ���Ŀ�ĵ�ϵ�� ;(5) ��֧��ɳ��ܿƵ�ϵ��;(6) ֧�ֻ���Ŀ���ܿƽ�Ԫ�ĵ�ϵ�ԣ��Ҹ��ܿƼ��ϵͳ������ϵΪ��( �����ܿ� + ( ���ܿ� + ( 䗛���ܿ� + ( ���ܿ� + ( ���ǻ��ܿ� + ( ţ���ܿ� + Ҷ����ܿ� ) + ( ͷ���ܿ� + ���ܿ� ))))))��
�����ؼ��� ��ֱ��Ŀ ;����������� ;�Ƚϻ����� ;����
����ֱ��Ŀ (Orthoptera) �����ڽ�֫������ (Arthropoda) ����� (Insecta)��Ŀǰȫ������������������ 27 941 �� (http://orthoptera.speciesfile.org/HomePage/ Orthoptera/HomePage.aspx)��ֱ��Ŀ���漸��ռ�������������ϳ��������������½����Ϣ�أ�����̬ϵͳ�о���������Ҫ�����ã��кܶ����ֱ���Ϊ����ѧ����̬ѧ�����ﻯѧ��������ѧ�Լ���������ѧ�������ģʽ���� [1-6]��Ŀǰ���������ձ���ͬ��ֱ��Ŀ��Ϊ����Ŀ(Caelifera)�����Ŀ(Ensifera)�����й���Ŀ�¸���Ԫ�Ļ�����Ȼ���ڷ��� [7]��ȷ����ֱ��Ŀ�������࣬�����˽�ֱ��Ŀ�������Դ������ϵͳ�����Լ���Ⱥ���ţ���ֱ��Ŀ������ء�����������Դ���þ�����Ҫ��ָ�����塣
�������������������Ϊ��״˫���Ĺ��۱պ� DNA ���ӣ�������Է�������С���ṹ���߿���������������Ա��ء�����������ʿ��Լ������ڼ��������������ĸϵ�Ŵ��ص㣬Ŀǰ�㷺Ӧ�����������ּ���������ϵͳ����ѧ����������ѧ�Լ���Ⱥ�Ŵ��ṹ��������о� [8-11]�������������Ÿ�ͨ������ķ�չ��������Ա�����Խ��Խ���ֱ��Ŀ������������������У�Ϊ�ӻ�����ˮƽ���о�ֱ��Ŀ��������ṩ�����õ�������
�������Ļ����˹�ȥ 22 ���ֱ��Ŀ������������������ijɹ��Լ�������ף��ܽᲢ�Ƚ��˻������С�������ɡ�������ʹ�á�����������ʡ��������ż�����ṹ��RNA ���������Ļ���������������������ֱ��Ŀϵͳ��������Ϊϵͳ�о�ֱ��Ŀ����ķ��༰ϵͳ���������˻������ϡ�
����1 ֱ��Ŀ��������������������״
����1995 �꣬Flook �� [12] �����˵�һ��ֱ��Ŀ������ɻ� (Locusta migratoria) �������������ȫ���С����Ų������IJ��Ϸ�չ������ 2017 �� 10 �� 13 �գ�GenBank ���ݿ��¼ 167 ��ֱ��Ŀ����ȫ��������������С��������Ի���Ŀ��������࣬���� 99 �֣��漰 9 ���ܿ� 22 ���� ;�������Ŀ�����ֹ��� 68 �֣��漰 7 ���ܿ� 12 ���ơ����б�ʵ���ҹ��ύ 53 �� (31.74%) ֱ��Ŀ��������������飬Ϊ�ؽ�ֱ��Ŀ������Ⱥ��ϵͳ������ϵ����ʾֱ��Ŀ����Ľ��������ṩ�˷ḻ�Ļ���������Դ ( ���� 1)����ȻĿǰ�ѻ��ȫ�������������������Ѹ���ֱ��Ŀ����������ܿƣ������Ӵ��ֱ��Ŀ���������ȣ���������ֻ�������������� Cooloolidae����� Mogoplistidae ��Ī��� Morabidae �Ȳ��ֿƼ���Ԫ���������֡�
����2 ֱ��Ŀ����������������������
����2.1��������������С
��������������������������������ƣ�Ŀǰ�Ѳ�õ�����ֱ��Ŀ����ȫ������������Ϊ�պϻ�״˫�� DNA�������� 13 �������ʱ������2 �� rRNA ����� 22 �� tRNA �������������������С��Ϊ Hemicharilaus monomorphus[13]������Ϊ 13 291 bp ;������Ϊ��ϸ��¶� (Phaneroptera gracilis)������Ϊ 18 255 bp��ֱ��Ŀ����������������С�ı仯����Ҫ�ܿ�����������������������ͳ���Ӱ�죬�����ɹ���� (Ruspolia dubia) [14] �Ŀ��������Ƚ��� 70 bp ;�������鳤�������ϸ��¶���˾���һ������Ϊ 1 548 bp �Ŀ��������� nad2 �� trnW ֮�仹����һ������Ϊ 1 830 bp �Ļ���������
����2.2����������������ṹ
�������ݻ���λ�ú�ת¼����ı仯������������������ſɷ�Ϊ��λ ( ����λ�øı䣬ת¼���� )����λ ( ����λ�ò��䣬ת¼����ı� ) �ͻ���ϴ�� ( �����������λ�� ( �� ) ��λ ) ������Ҫ������ [15]��Ŀǰ�Ѳ�������������ֱ��Ŀ������� 8 ��������������������� ���� 54 ��ֱ��Ŀ������ͼ 1-1 ��ʾ�ĵ��͵Ľ�֫����������������鷽ʽ���У�������������Ŀ���� (50 �� ) �ͼ������Ļ���Ŀ���� (4 �� ) ;�� 90 ��ֱ��Ŀ����������������� trnK �� trnD ��������λ������ͼ 1-2 ��ʾ�� trnD-trnK ��ʽ���� ( Ҳ�� KD ���� )��������㷺�����ڳ������ܿƺ����ܿƵı�ɫ���� (Erianthus versicolor) [13] ����Ļ���Ŀ�����У������Ŀ������� KD ��������;�� 9 ��ֱ��Ŀ����������������� trnN��trnS �� trnE ����������ϴ�ƣ�����ͼ 1-3 ��ʾ�� (-trnE)-(-trnS)-(-trnN)��(-trnE)- (trnS)-(-trnN) �� (-trnE)-( trnS)-(trnN) ���ַ�ʽ���ڣ���������������е����ܿ��� ;�� 3 ��ֱ��Ŀ����������������� trnI��trnQ �� trnM ����������ϴ�ƣ�����ͼ 1-4 ��ʾ�� trnM-trnI-(-trnQ) ��ʽ���ڣ����������������Ҷ�x�ǿ� (Pseudophyllinae) �� ;�� 3 ��ֱ��Ŀ����������������� AT ��������trnI��trnQ��trnM �� nd2 ����������ϴ�ƣ�����ͼ 1-5 ��ʾ�� trnI-trnM-nd2-AT-(-trnQ) ��ʽ���ڣ��������������¶�Ƶĸ�����¶� (Holochlora fruhstorferi)�����ѻ���¶� (Sinochlora longifissa) [16]���Ĵ�����¶� (Sinochlora szechwanensis) �� ;���Ƶ� Cyphoderris monstrosa[6] ������������� trnA �� trnR ��������λ������ͼ 1-6 ��ʾ�� trnR-trnA ��ʽ���� ;�˹�Ƶ������ (Lipotactes tripyrga) ������������� trnG��nd3��trnA��trnR��trnN �� trnS ����������ϴ�ƣ�����ͼ 1-7 ��ʾ�� trnRtrnS-trnA-trnN-trnG-nd3 �ķ�ʽ���� ;¶�Ƶ��۳�ھ�¶� (Ruidocollaris obscura) [17] ������������� AT ������ͨ������ϴ�Ƴ����� trnY �� cox1 ֮�䣬����ͼ 1-8 ��ʾ�� (-trnY)-AT-cox1 �ķ�ʽ���ڡ�
���������� 4 ���Ѳ�ֱ��Ŀ�����д�������������������ṹ���ֱ�Ϊͷ�ȿƵĽ����� (Mekongiella kingdoni) [18] ȱ�� trnR ������ֺ�Ƶ� Comicus campestris[6] ȱ�� trnM �����ǻȿƵ� Locusta migratoria migratoria ���շ�Ƶ� Troglophilus neglectus[19] �� cox1 �� cox2 ������� 2 �� trnL��������˵�������Ĥ��Ŀ�ȸ��³�Ŀ���� [20]��ֱ��Ŀ��������������������¼���Ա��أ����˻���Ŀ�� KD ���ţ����� 23 ��ֱ��Ŀ����������������������ṹ���������š�Ŀǰ���Ի�����������Ľ�����Ҫ�и��Ʒ������ʧģ�͡��������ɾ��ģ�͡������Լ��� tRNA ���������ʼ����ĸ��� 4 �ֽ��� [21]������Щģ���У��������ɾ��ģ���Ѿ���ò���֤�ݵ�֧�֣���������Ȼû��һ��������ģ�Ϳ��Խ������е��������� [22]����ˣ����ڸ߶����ŵ����������������ö��ģ�ͽ��ͣ����磬���ڽ����� KD ��������Ļ���Ŀ�����ø������ɾ��ģ�ͽ��ͣ������ڷ����˻���ϴ�Ƶ��������͵������������������Ҫ��ϸ���ɾ��ģ�ͼ�����ģ������ͬ���͡�
����2.3����������������ʹ�����
�����Ѳ��ֱ��Ŀ����ȫ�����������������Ե� AT ƫ���� ( ���� 2)��ƽ�� A+T ����Ϊ 73.31%������ A+T ������ߵ�Ϊ��Ƶ� Paramastax nigra[13] (78.00%)�����Ϊ������� (Gampsocleis gratiosa) [23] (65.30%)��
������ֱ��Ŀ��Ŀ���ܿƽ�Ԫ��ȫ������������ƽ�� AT ���������ȽϷ��� ��2 ����Ŀ�У�����Ŀ�� A+T ���� (74.24%) �������Ŀ (71.95%) ;����Ŀ�� 9 ���ܿ��У�Ҷ����ܿƵ� A+T ������� (78.00%)���������ܿ���� (68.80%) ;���Ŀ�� 7 ���ܿ��У�������ܿ� A+T ������� (75.00%)����ԭ��ܿ���� (69.45%)��һ����˵��ͬһ�Ƽ���Ԫ�е����ּ������������С�������ڻ���Ŀ�У� A+T �����ڲ������������Ľ��ǻȿ� (SE=0.12) ���ȿ� (SE=0.15) �г��ֳ����Ե���̬ ;������������ˣ����������Ŀ�У����������������˹�� (SE=0.48)��¶�� (SE=0.33) ����� Gryllidae (SE=0.63) �У���ͬ�����ּ� AT ��������ϴ� ( ͼ 2)�����Ƶ�����Ҳ�����ڰ��Ŀ�� [22]��
���������ձ���֯�� (Mecopoda niponensis) [24]��������¶� (Holochlora fruhstorferi) [11]���Ĵ�����¶� (Sinochlora szechwanensis)���ڽ���¶� (Phaneroptera nigroantennata)��Tridactylus sp. �� Mirhipipteryx andensis[6] �⣬�����Ѳ�ֱ��Ŀ����������ȫ������� 4 �ּ��������Ϊ As>Ts>Cs>Gs��ͬʱ�����ֳ���ǿ�� A ����� C ���ƫ�� (AT-Skew = 0.11�� GC-Skew = -0.21)��ֱ��Ŀ�����������������ֳ��ļ�����ƫ�������ڽ����Ĺ�����ͻ�䡢��Ȼѡ������Ŵ�Ư�䡢ˮƽ����ת�ƺͻ�����ṹ�ȶ������������أ����������������������A>T��C>G��������ƫ����A��C���ּ��[25]������һ�������������������� AT ��ƫ��� GC ��ƫ����ص� [26]��
������ֱ��Ŀ������Ŀ���ܿƽ�Ԫ��ȫ���������������ɵķ����ȽϷ��� ( ͼ 3) ������Ŀ�����Ŀ�ļ��ƫб�����Ϻ��������������������ձ���� ;����Ŀ�ļ�����ƫб��ǿ�� (AT skew = 0.15, GC skew = -0.18)�����Ŀ������ AT ��������Ϊ�ӽ� (AT skew = 0.05, GC skew = -0.27)������Ŀ 9 ���ܿƵļ��ƫб�������ձ���ɣ��������ܿ�������AT��������Ϊ�ӽ�(AT skew = 0.02)�⣬���� 8 ���ܿ� AT ƫб��Ծ���ǿ�ң�AT ƫб��ΧΪ0.11~0.23;����䗛���ܿƵļ�����ƫб��ǿ(AT skew = 0.23, GC skew = -0.23)�������ܿƼ�����ƫб���� (AT skew = 0.11, GC skew = -0.14)�����Ŀ 7 ���ܿƵļ��ƫб�������ձ���ɣ��� AT ƫб������ڻ���Ŀ��AT ƫб��ΧΪ 0.02~0.09 ;����ԭ��ܿƵļ�����ƫб��ǿ�� (AT skew = 0.09, GC skew = -0.29)�����ܿ� (AT skew = 0.08, GC skew = -0.28) ��֮����������ܿ� (AT skew = 0.02, GC skew = -0.24) ������
�����ֱ�ͳ�� 167 ���Ѳ�ֱ��Ŀ��������������鵰���ʱ�������������ʹ����������� 2 ����Ŀ�����ͬ�������ӵ�ʹ��Ƶ�� (relative synonymous codon usage, RSCU)��ͳ�ƽ����ͼ 4 ��ʾ �������������У�UUA(L) �ڻ���Ŀ�����Ŀ�е� RSCU ֵ��Ϊ��ߣ��ֱ�Ϊ 2.91 �� 2.74 ;GCG(A) �ڻ���Ŀ�����Ŀ�е� RSCU ֵ��Ϊ��ͣ��ֱ�Ϊ 0.11 �� 0.22 ;UGG(W) �� AUG(M) �ڻ���Ŀ�����Ŀ�о���ƫ���� (RSCU = 1) ;������ʹ��ƫ�����������ӵ���λ��� AT ƫ���Գ��ֳ�һ������ԣ�����λ��Ϊ A �� U ��������ʹ��Ƶ���ձ�ϸ� ;ͬһ�������ӵ� RSCU ֵ��ֱ��Ŀ 2 ����Ŀ��������������졣
����2.4��ֱ��Ŀ���������嵰���ʱ������������
����ֱ��Ŀ����� 13 �������嵰���ʱ������� A+T �����������Ե������������������(����2)��
����������ƣ������ʱ������ĵ�����λ�� A+T ����Զ���������ӵ�һ����λ�㣬����������������Ľ��ǻȿ������ӵĵ���λ�� A+T �����ߴ� 76.38%���������ӵ�һλ��͵ڶ�λ���Ϊ 73.85% �� 72.13%��
����ͨ���Ƚϣ�ֱ��Ŀ������ʼ�����Ӿ��������ص� ( ��ͼ 1) ��atp6��cox2��cytB��nd3 �� nd4 ����ʼ�����Ϊ�������������� ATN (ATA��ATT��ATC�� ATG) ;cox3��atp8��nd1��nd2��nd4��nd5 �� nd6 ����ʼ�����ӳ��˱������������� ATN ���⣬���н��������ʼ�������� GTG��TTG��AGT��TTA�� CTG �� CCT ;���� cox1 �������ʼ��������Ϊ���ӣ����������������� ATN ��Ϊ��ʼ�����ӣ��� cox1 ʹ���˷DZ������ӣ����� CCG��CAA�� ATT��TTA��ACC��ACG �ȣ����ⲻ��������������� ATGA[27]��GTGA[13]��ATAA[12] �ȶ��ǿ��ܵ� cox1 ����ʼ�����ӡ���Щ�������ʼ�����ӿ��Լ�С����������ͬʱ�ֱ������ڻ������ص���ͬʱ����ת¼��Ϊ mRNA �� RNA �༭ת������������ʼ�����Ӳ���������ķ������ [22, 28]��
��������ʼ�����Ӳ�ͬ��ֱ��Ŀ��������ֹ�����ӷ�����ֳ��ϸߵ�һ���� ( ��ͼ 2) ��atp8 ����ֹ������Ϊ���͵����������� TAA��TAG ;���� 12 �ֵ����ʱ���������ֹ�����ӳ������������������� TAA �� TAG ֮�⣬����������������ֹ�������� TA �� T�����ֲ���������ֹ�������ں��������������������кܳ������о����Ʋ������ת¼��ͨ����������������γ���������ֹ�����ӽ������ת¼��ֹ [29]��
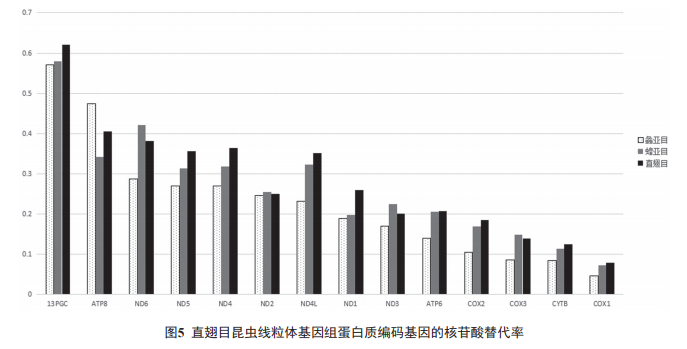
��������������������鵰�ױ����������ܵ���л�м������ձ䣬����ں˻�������нϸߵĽ������� [30]����ֱ��Ŀ 2 ����Ŀ�����������ĵ����ʱ������������� ( �÷�ͬ���������ͬ������ʵı��� Ka/Ks ����ʾ�������� ) ���жԱȣ����������ͼ 5 ��ʾ ����ͬ�ĵ����ʱ������Ľ������ʲ�ͬ������ atp8 �������������죬�� cox1 ��Ϊ���أ�13 �ֵ����ʱ��������ֱ��Ŀ�еĽ�������˳��Ϊ atp8 > nd6 > nd4 > nd5 > nd4L > nd1 > nd2 > atp6 > nd3 > cox2 > cox3 > cytB > cox1 ;��ͬ�ĵ����ʱ�������ڲ�ͬ����Ⱥ��ͬ�����������IJ��죬�� atp8 �������� 12 �������ʱ�������ڻ���Ŀ�еĽ������ʾ��������Ŀ ;13 �������ʱ������� Ka/Ks ��С�� 1�����ֳ����� ( �� ) ѡ����������ͬ��ͻ�����Ϊ���Ʊ���̭�ˡ�
����2.5 tRNA��rRNA�ص�
�����Ѳ��ֱ��Ŀ����������������У���� trnSAGN ȱ�� DHU �ۣ�ȱʧ DHU �ۺ�� trnSAGN �Կ��γɵ� L �������ṹ��ά�� CCA ���ܱ��뷴�����Ӽ�ľ��� [31] ;���� 21 �� tRNA �����۵��γɵ��͵���Ҷ�ݽṹ����ȻҲ�����⣬��ɽ�Ļ� (Sinopodisma houshana) �� trnP ֻ�� 58 bp���ɱ价�� TΨC �۾������� [32]��tRNA ���γ���Ҷ�ݽṹ������ʱ�ᷢ��һЩ���䣬��Ҫ�� G-U ���䡣���������������о��ձ���ڣ��ڶ�֩�� (Araneida) ������ tRNA ���о��з��֣���������ļ������ͨ��ת¼��༭�ָ�������ԣ��Ӷ��Ʋ������� tRNA ����ȱʧ DHU �ۻ� T �۶����������ܵ���ʹӰ�첻�� [33]�����⣬ֱ��Ŀ���� tRNA Я���ķ������ӷdz��̶�����δ������������ķ������ӡ�
�������֪ʶ�Ƽ���������ʵ�����ķ���
����rRNA ������� rrnL ���� (lrRNA �� 16S rRNA)�� rrnS ���� (srRNA �� 12S rRNA)�����С��λ����Ա��أ��Ҽ����ɱ��ֳ����Ե� AT ƫ�á� rrnL ����Ķ����ṹ���� 6 ���ṹ�� (I��II��III�� IV��V �� VI)�����нṹ�� III �ڽ�֫�������������������ȱʧ�ģ��ṹ�� IV �� V ���س̶Ƚϸ� ; rrnS ����Ķ����ṹ�����ĸ��ṹ�� (I��II��III �� IV)��ͨ���ṹ�� I �� II ����ϴ� III �� IV �Ƚϱ��� [34]��ֱ��Ŀ����������������� rrnL ����λ�� trnLCUN �� trnV ֮�䣬ƽ������Ϊ 1 317 bp���������Ϊ���dz�� (Calliptamus abbreviatus)������Ϊ 1 555 bp ;��̵�Ϊŷ������ (Gryllotalpa pluvialis)������Ϊ 1 236 bp[19]��ֱ��Ŀ��������������� rrnS ����ij���ԶС�� rrnL ����ƽ�����Ƚ�Ϊ 800 bp���������Ϊ���Ƶ� Trigonidium sjostedti������Ϊ 1 004 bp[35]������̵�Ϊ��ɫ��� (Phlaeoba tenebrosa)�����Ƚ�Ϊ 212 bp��
����2.6���������ص�
����ֱ��Ŀ���������λ����Ա��أ�ͨ��λ�� rrnS �� trnI ֮�䣬��Ҳ�����⣬���縵����¶���ѻ���¶���Ĵ�����¶�Ŀ�����λ�� nd2 �� trnQ ֮�䣬��ھ�¶�Ŀ�����λ�� trnY �� cox1 ֮�� ;���Ȳ���ܴ�������̵�Ϊ�ɹ���� [14]������ 70 bp�������Ϊ���ѻ���¶� [16]������������Ϊ 3 122 bp����ͬ���ּ�������ij��ȱ仯��Ҫ��߶��ظ������е������ʹ�С��أ���Щ�ظ����пɽ�һ���γɼķ��нṹ���Ϊ���ӵĶ����ṹ���谭���ƵĽ��̡����о����������ܲ�ͬ���������� N ���ĸ������λ�ò�ͬ����һЩȫ��̬������������д��ڵ� T-strech (ploy-T > 10 bp) �����븴����ʼʶ���йأ������ְ��̬���������������鲢���� T-strech �ṹ [36]����ֱ��Ŀ�У��ֻ���Ŀ�Լ����Ŀ�����ܿƺ������ܿ���������������������Ȼû�� T-strech �ṹ�����о��������Ӧλ�÷��� ploy-T �Ľṹ����� C ��ϣ����γ�����Ӧ�ľ����ṹ��������ɻ�(Locusta migratoria) [12]������Ļ� (Sinopodisma tsinlingensis) [37]��ɮñ��� (Phlaeoba infumata) [37]��С��� (Traulia minuta) [37] �� ;�����Ŀ���˹�ܿƼ�������Ԥ������ľ����ṹ [34]�����о������˹�ܿƲ������ֵ������������ N ���Ϸ����� T-strech �ṹ������Ħ���˹ (Anabrus simplex) [38]��������� (Deracantha onos) [39]��������� (Gampsocleis gratiosa) [38] �ͺڽ���¶� (Phaneroptera nigroantennata) [37]���ܶ���֮����������������������г��Ⱥͽṹ����ܴ�——�������ߣ��ԡ���1,2����ѩ��1 ���ơ�ԭ1 *